陆剑研究组合作开发免疫印迹数学模型DynaVac,定量指导疫苗更新与接种策略
2026年4月16日,基因功能研究与操控全国重点实验室、北京大学生命科学学院陆剑研究组联合云南大学张子杰研究组,在Cell Reports发表题为“Mapping immune imprinting zones enables predictive vaccination optimization”的研究论文。该研究聚焦病毒持续变异背景下疫苗更新面临的核心难题——免疫印记(immune imprinting),构建了一个可定量预测抗体应答的动力学模型DynaVac。研究整合了小鼠实验及人体临床试验中的中和抗体数据,系统刻画了抗原距离、加强剂量和接种时程等因素如何共同塑造免疫印记效应,并提出了“保护区—陷阱区—突破区”三分区框架,为变异株疫苗的更新与加强策略优化提供了预测性理论工具。
RNA病毒因复制速度快、突变率高、演化迅速等特征,使SARS-CoV-2等病毒在流行过程中不断产生逃逸既往免疫的新变异株。在此背景下,既往感染或疫苗接种所形成的免疫记忆,在再次暴露于新抗原时会被优先唤起,进而影响针对新变异株的抗体应答,这一现象即免疫印记。免疫印记既可保留对既往毒株的保护,也可能抑制针对新变异株的从头应答,成为变异疫苗优化的关键障碍。尽管大量研究已观察到该现象,但由于免疫史、疫苗平台和变异株之间的复杂组合,仅凭经验难以判断何种接种方案能够有效更新免疫记忆以建立对新变异株的保护。
陆剑团队长期聚焦RNA病毒进化与疫苗优化研究,前期已率先提出SARS-CoV-2的S/L双谱系划分并构建分层次谱系系统,揭示了病毒演化与传播的基本轨迹(Tang et al.,National Science Review, 2020);同时从密码子演化角度提出mRNA疫苗序列优化策略(Wu et al.,Advanced Science, 2023);并建立SIRSVIDE模型刻画病毒传播、免疫逃逸与演化动态(Jin et al.,hLife)。在此基础之上,本研究进一步建立了基于常微分方程的动力学模型DynaVac,从机制层面同时刻画抗原、初始B细胞、记忆B细胞和抗体之间的动态关系,实现了对复杂序贯接种条件下体液免疫反应的定量模拟。
在模型构建过程中,研究首先将体液免疫反应划分为初次免疫和加强免疫两个阶段。在初次免疫阶段,模型描述抗原产生、初始B细胞活化与成熟、记忆B细胞形成以及抗体分泌等关键过程(图1A、B);在加强免疫阶段,进一步引入不同变异株之间的交叉中和关系,以及既有记忆B细胞与初始B细胞对新抗原的竞争(图1C、D)。相较于传统高复杂度的单细胞水平模拟,DynaVac采用群体层面的平均化建模策略,在保留主要免疫学机制的同时大幅降低计算量,因而能够支持大规模参数拟合和多种接种方案的前瞻性模拟。
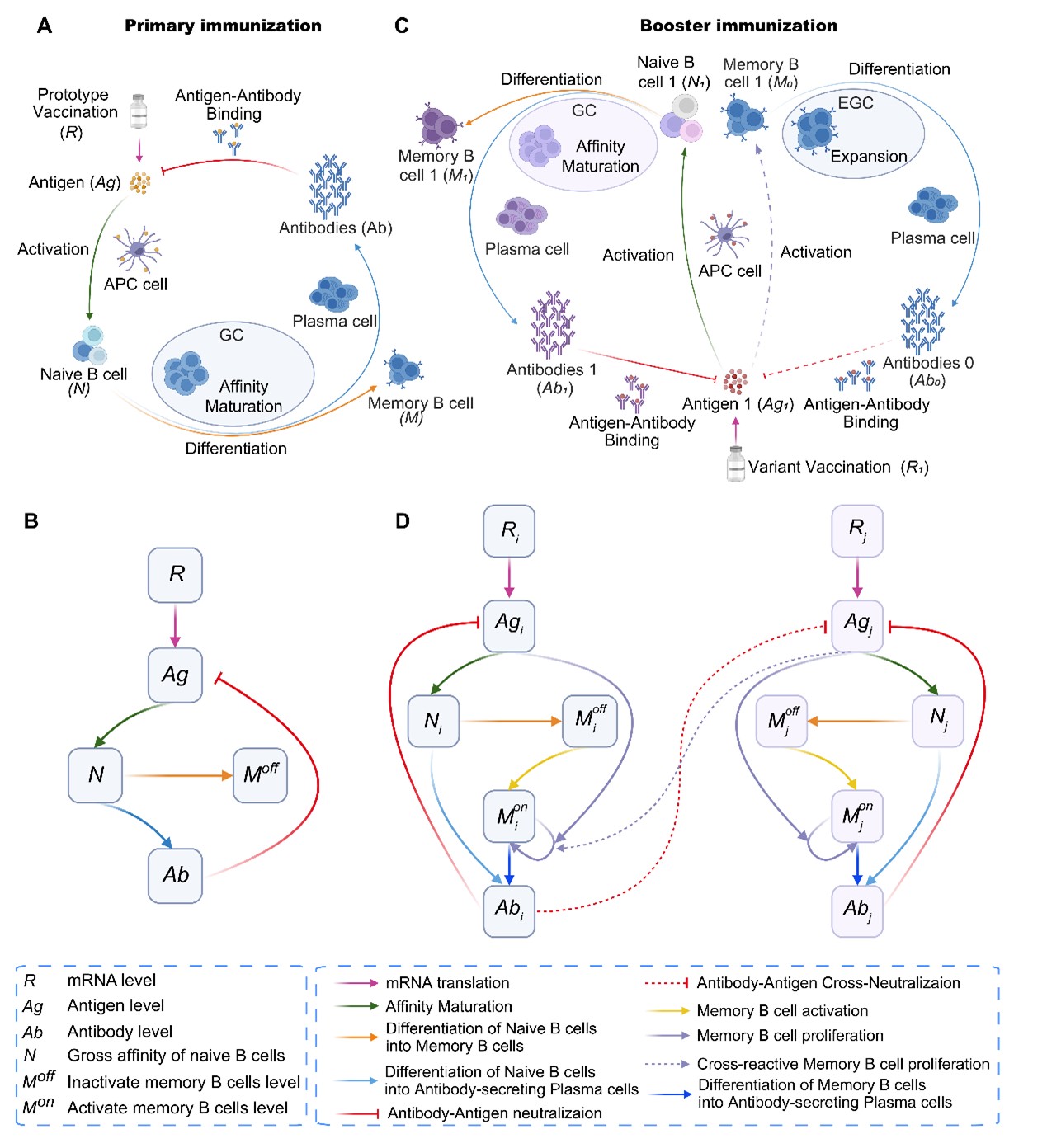
图1:DynaVac模型构建与免疫应答动力学框架。A. 初次免疫过程中,mRNA来源抗原诱导初始B细胞活化、亲和力成熟、记忆B细胞形成及抗体产生的示意图;B. 初次免疫阶段关键免疫变量之间的抽象动力学关系;C. 异源加强免疫时,既有记忆B细胞与新激活的初始B细胞共同参与应答的示意图;D. 多变异抗原条件下交叉中和与交叉记忆激活的整体建模框架。
为验证模型性能,研究团队设计了涵盖原始株及多个主要SARS-CoV-2血清型变异株的29种小鼠免疫程序,并结合已发表的数据集以及人体临床试验数据,对模型进行了系统参数化与交叉验证。结果显示,DynaVac能够对不同接种程序下、针对多种病毒变异株的中和抗体反应进行定量预测,在小鼠和人体临床试验数据中均表现出良好的预测准确性(图2)。这表明,该模型不仅能够解释已有实验观察,更具备对不同免疫背景、疫苗平台及接种策略下抗体应答结果进行前瞻性预测的能力。
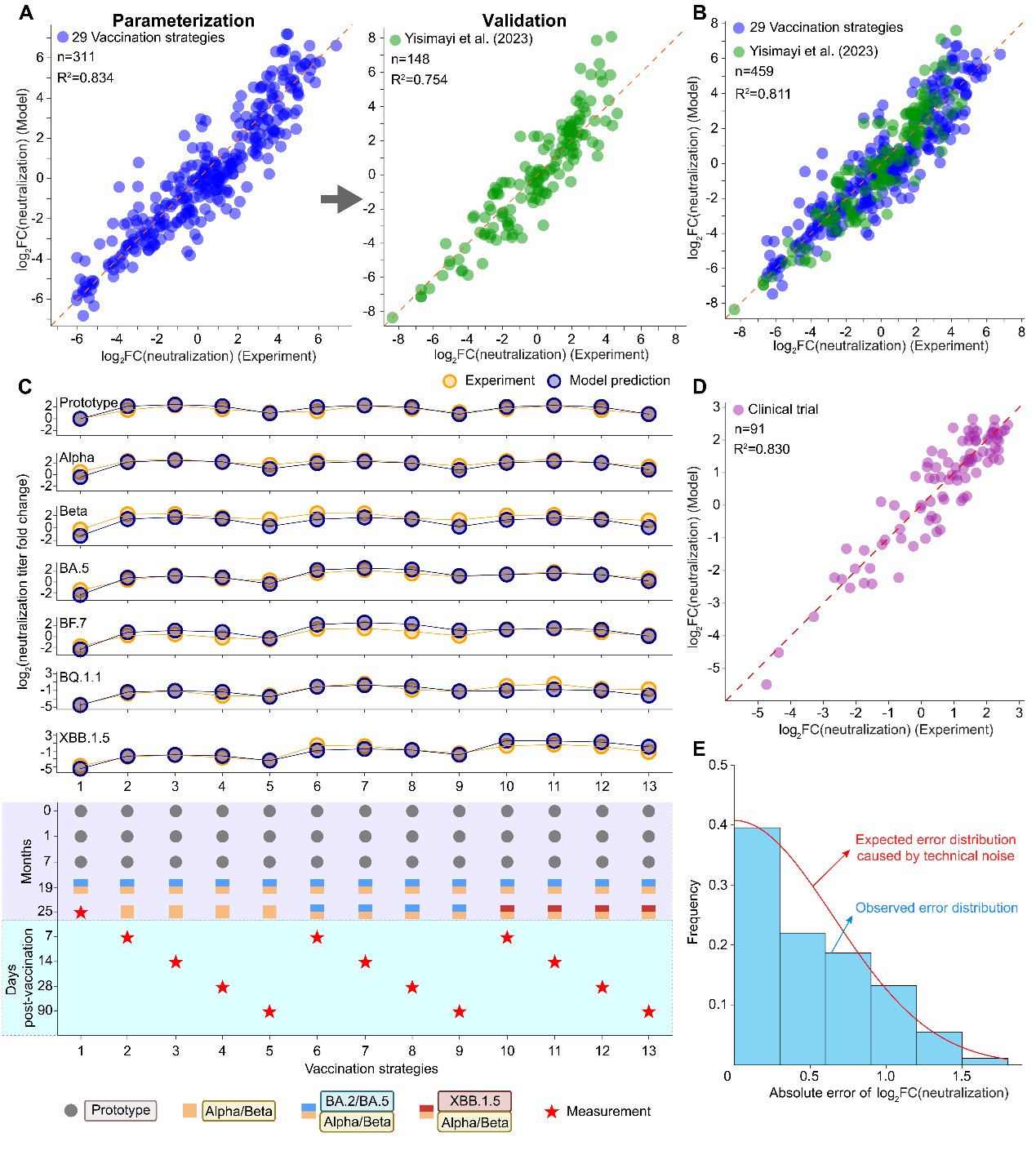
图2:DynaVac模型在小鼠和人体数据中的参数化与验证。A. 模型在本研究小鼠数据集及独立小鼠数据集中的交叉验证结果;B. 基于合并小鼠数据集参数化后的整体预测性能;C. 人体临床试验中不同时间点、不同接种方案下的实验中和抗体滴度与模型拟合结果比较;D-E. 模型在人群数据中的整体拟合精度及误差分布。
在此基础上,研究进一步利用DynaVac对不同抗原距离下的加强免疫过程进行了系统模拟(图3A)。结果发现,初始免疫株与加强免疫株之间的抗原距离是决定免疫印记效应走向的核心参数。当两者抗原距离较近时,原有抗体能够有效交叉中和新抗原,形成“免疫印记保护区”;当抗原距离处于中间范围时,既有记忆仍足以压制新生应答,但交叉中和又不足以提供有效保护,从而形成“免疫印记陷阱区”;当抗原距离足够远时,原有记忆对新抗原的压制明显减弱,新的变异株特异性应答得以建立,进入“免疫印记突破区”。这一连续的免疫景观,为理解为何某些变异株加强针效果有限、而另一些则能有效刷新免疫记忆,提供了统一的解释框架(图3C)。
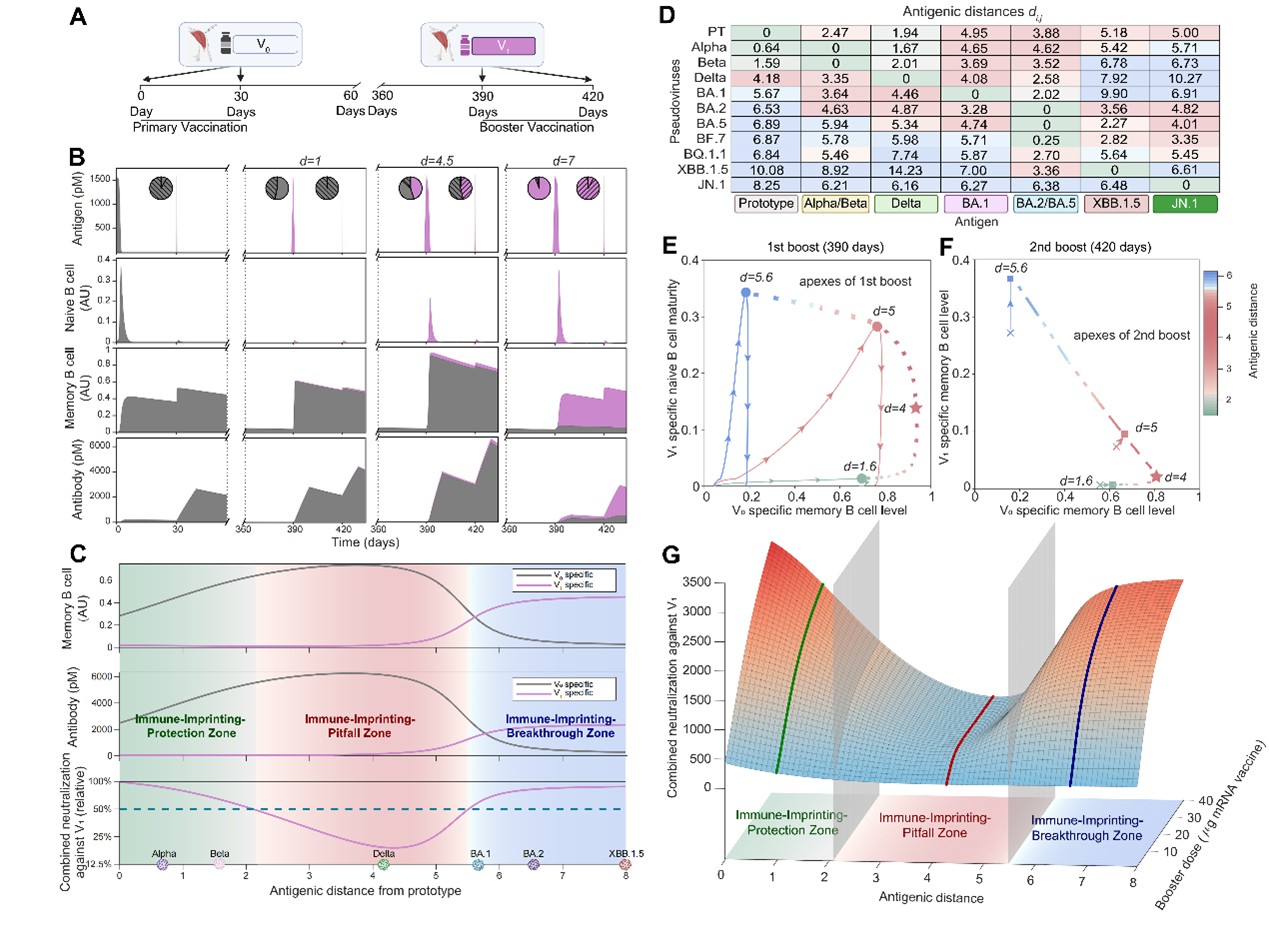
图3:抗原距离决定免疫印记的“保护区—陷阱区—突破区”。A. 模拟的接种流程:两剂原始株疫苗后,再进行两剂变异株加强;B. 在不同抗原距离条件下,抗原、初始B细胞、记忆B细胞与抗体的动态变化;C. 变异株中和能力随抗原距离变化呈U形分布,据此划分保护区、陷阱区和突破区;D. 主要SARS-CoV-2变异株与原始株之间的实验抗原距离及其在三类分区中的对应位置;E. 第一次加强免疫过程中,变异株特异性初始B细胞与原始株特异性记忆B细胞之间的竞争轨迹,展示不同免疫印记分区内早期应答主导群体的变化;F. 第二次加强免疫过程中,变异株特异性记忆B细胞与原始株特异性记忆B细胞之间的竞争轨迹,反映免疫特异性在连续加强后的重塑方向。G. 加强剂量与抗原距离共同塑造对变异株中和能力的变化趋势。
进一步分析表明,免疫印记本质上源自既有记忆B细胞与新激活初始B细胞之间的动态竞争。在初次加强免疫期间,哪一类细胞占据主导地位,决定了后续抗体应答的特异性方向。在保护区内,原始株记忆B细胞几乎完全主导应答;在陷阱区内,虽然初始B细胞有所参与,但仍难以扭转既有的免疫偏向;而在突破区内,变异株特异性初始B细胞及其后续形成的记忆B细胞得以逐步占据优势,从而推动免疫特异性的更新(图3E、F)。值得注意的是,单纯提高加强剂量并不能有效摆脱陷阱区中的免疫印记抑制,说明“加大剂量”并非普适的解决方案 (图3G)。
研究还系统评估了初免剂量和初免—加强间隔对免疫印记的影响。模拟结果显示,较高的初免剂量会强化原始株免疫记忆,从而扩大陷阱区范围;相反,较低的初免剂量有助于缩小陷阱区、扩大突破区。与此同时,延长初免与加强之间的时间间隔可使原有记忆逐渐衰减,也有利于减轻免疫印记对变异株应答的压制。这表明,除抗原成分外,剂量与时间窗口同样是决定后续疫苗更新效果的重要调控因素(图4)。
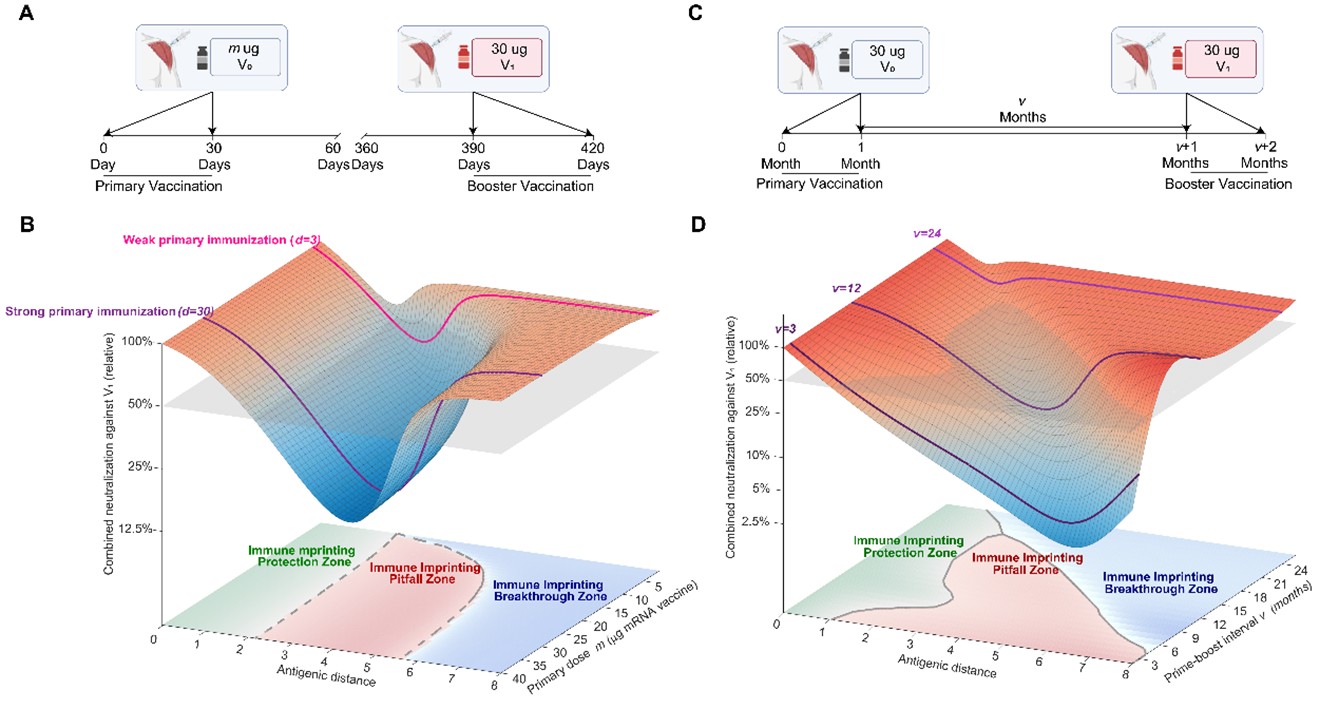
图4:初免剂量和接种间隔共同塑造免疫印记效应。A-B. 初免剂量与抗原距离共同塑造加强接种对变异株中和能力,并影响免疫印记保护区、陷阱区和突破区的分布范围。C-D. 初免—加强间隔与抗原距离共同塑造加强接种对变异株中和能力,并影响免疫印记保护区、陷阱区和突破区的分布范围。
综上所述,该研究建立了一个可定量预测复杂序贯接种免疫结果的动力学框架DynaVac,提出并系统定义了免疫印记的“保护区—陷阱区—突破区”三分区模型,揭示了抗原距离、接种剂量和免疫时程如何共同塑造变异株疫苗的应答方向。研究不仅为理解SARS-CoV-2持续变异背景下的疫苗更新难题提供了新的理论框架,也为未来流感、HIV等快速演化病原体的疫苗优化提供了可推广的建模思路。更重要的是,这项工作推动疫苗设计由经验驱动走向预测驱动,为在不断变化的病毒抗原景观中实现更精准的疫苗更新开辟了新路径。
云南大学生物交叉技术创新中心杨薇助理研究员,北京大学生命科学学院博士生靳垲淳和云南疫苗实验室mRNA BU负责人李薇为论文的共同第一作者。云南大学生物交叉技术创新中心张子杰研究员和北京大学生命科学学院陆剑教授为本论文的共同通讯作者。云南大学丁志兴,喻厚泽,周祖密,苏炜,朱潇,马旭普,万娜,邢蕾,陈燕丽,余泫静,北京大学生命科学学院博士后唐小鹿,复旦大学林金钟教授,云南大学信息学院李浩教授也对本论文做出了重要贡献。该工作得到了科技部国家重点研发计划、广州国家实验室专项项目、云南省西南联合研究生院科技项目、北京市自然科学基金、国家自然科学基金、云南省基础研究计划、云南疫苗实验室研发项目等机构的支持,并依托北京大学高性能计算平台、北京大学生命科学中心高性能计算平台和国家生物医学成像中心生物医学计算平台完成相关研究工作。
- 钟上威课题组与合作者揭示泛素连接酶UPL3调控光诱导种子萌发的新机制2026.06.11
- 魏文胜团队联合合作者在RNA编辑治疗杜氏肌营养不良症研究中取得重要进展2026.06.11
- 魏文胜团队开发新一代RNA编辑技术LEAPER 3.02026.06.11
- 探秘前沿科技,播种科学梦想——2026年全国科技周科普活动圆满举办2026.06.09
- 秦跟基研究组揭示叶片衰老的关键调控机制2026.05.26
- 重塑时间韧性:王伟团队首次系统解析植物生物钟在极端环境中的适应性调控新范式2026.05.13
- 胡家志课题组构建新型高频突变平台实现抗体高效定向进化2026.05.12
- 钟声团队揭示被子植物配子互作的新机制2026.05.08
- 王伟课题组与合作者揭示大豆孢囊线虫劫持大豆生物钟的新机制2026.05.08
- 邓伍兰课题组首创 SMLDM 显微技术:单帧解析单分子定位与扩散,突破活细胞动态成像瓶颈2026.04.30
- 张蔚课题组与合作者揭示潮间带蜘蛛跨越陆海界限极端生存的基因组策略2026.04.28
- 陆剑研究组合作开发免疫印迹数学模型DynaVac,定量指导疫苗更新与接种策略2026.04.20
- 汪阳明团队开发REDDIT技术,实现增强子RNA与多类非编码RNA转录的高灵敏动态监测2026.04.20
- 陆剑课题组揭示果蝇上游开放读码框的演化规律与补偿机制2026.03.27
- 魏文胜课题组受邀综述:利用碱基与引导编辑器实现疾病相关变异的大规模解码2026.03.23
- 焦雨铃研究组构建禾本科花序发育数学模型,指导小麦高产基因挖掘2026.03.16
- 何爱彬团队与合作者开发新一代表观液体活检平台实现疾病组织器官精准溯源2026.03.05
- 陈雪梅教授课题组揭示新型RNA帽子修饰的跨物种分布与动态调控2026.03.05
- 肖俊宇团队与赵明辉、谭颖团队合作揭示补体系统C3转化酶组装及识别底物C3的分子机制2026.02.21
- 钟声课题组鉴定到拟南芥隔膜处“阻止多花粉管穿出”受体复合物的新组分2026.02.11
- 钟声研究员与瞿礼嘉院士合作揭示被子植物两次受精事件对受精补偿效率的关键作用2026.02.11
- 陈雪梅研究组揭示RdDM通路在作物免疫调控中的新功能,为大豆抗病改良提供了潜在的新思路、新靶点2026.02.10
- 秦跟基课题组揭示调控种子大小的新机制2026.02.10
- 罗述金课题组古DNA研究揭示:日本晚更新世并无虎,而为洞狮2026.01.27
- 张蔚课题组与合作者揭秘蜘蛛纺器起源2026.01.15
- 伊成器研究组报道AIM平台,突破“单点编辑”限制,实现RNA多位点、多功能精准操控2026.01.07
- 田望课题组发现植物抗逆“全能选手”,为提高植物“钙营养”和复合逆境抗性提供新概念2026.01.06
- 张蔚课题组与合作者揭示红珠凤蝶马兜铃酸抗性机制2026.01.05
- 刘君团队开发FOCAS平台实现m6A修饰位点的全转录组功能解析2026.01.05
- 肖俊宇研究组揭示FcRL5受体识别 IgG的分子机制2026.01.02
- 王伟课题组报道应激颗粒实现胞内氧化还原异质性的新功能:细胞抗氧化的“智能调控枢纽”2025.12.22
- 李晟课题组与合作者揭示保护地缓解人类对兽类行为干扰的关键作用2025.12.15
- 姚蒙研究组系统评估无脊椎动物宏条形码引物,助力eDNA生物多样性研究2025.12.12
- 朱丹萌课题组与合作者揭示PICKLE调控染色质状态与基因转录的分子机理2025.12.09
- 季雄团队提出FeaSion策略揭示RNA聚合酶磷酸化的特征调控与功能2025.12.01
- 罗述金课题组古DNA研究揭示:豹猫与人共栖3500年直至汉末,家猫经丝绸之路唐代方始传入2025.11.28
- 白洋研究员入选第三期 “新基石研究员项目”资助名单2025.11.26
- 实验室主任瞿礼嘉教授2025年新当选中国科学院院士2025.11.25
- 汪阳明团队与西湖大学卢培龙团队利用人工智能辅助蛋白质设计实现精准线粒体DNA碱基编辑2025.11.18
- 肖俊宇和高宁团队联合发表多聚抗体设计策略2025.11.07
- 赵进东课题组应邀撰写藻胆体的结构和能量传递机制长文综述2025.10.28
- 郑晓峰课题组揭示代谢酶ALDOA入核激活NF-κB信号通路驱动胰腺癌进展2025.10.23
- 陆剑课题组揭示果蝇饥饿耐受的遗传架构和性别差异2025.09.29
- 遇赫课题组揭示中国家猪的起源传播与人群互动过程2025.09.26
- 季雄团队揭示INO80/SWR复合物通过BRD2和染色质景观调控Pol II转录起始2025.09.22
- 刘启昆课题组与合作者在Current Opinion in Plant Biology发文总结植物再生领域表观遗传调控的研究进展2025.09.18
- 钟上威课题组受邀发表植物光信号传导Tansley长文综述2025.09.16
- 姚蒙课题组研发新型空气eDNA采集器高效检测动植物多样性2025.09.11
- 焦雨铃研究组克隆了小麦粒重新基因,服务高产育种2025.09.02
- 魏文胜团队与合作者联合开发新型通用型CAR-T 疗法治疗血液系统恶性肿瘤2025.08.22
- 魏文胜团队开发RNA环化新方法2025.08.12
- 汪阳明团队创新双组学技术MAPIT-seq:在单细胞水平同时绘制RNA结合蛋白作用图谱与基因表达图谱的新利器2025.08.11
- 周岳课题组揭示拟南芥PRC1介导的H2Aub loop的形成机制及功能2025.08.11
- 贺新强课题组与周岳课题组合作揭示PRC2调控维管组织模式建立机制2025.08.11
- 苏晓东课题组揭示清道夫受体CD163识别底物的分子机制2025.07.23
- 陈雪梅课题组系统阐释植物miRNA成熟与功能调控机制2025.07.23
- 苏晓东课题组与合作者揭示AAV与新型受体(AAVR2)结合的分子机制2025.07.15
- 合成植物基因组2025.07.09
- 伊成器课题组与合作者建立RNA"暗码",升级生命语言2025.07.04
- 魏文胜团队系统揭秘人类基因组中“沉默突变”的功能性“低语”2025.06.26
- 肖俊宇团队与张永辉团队合作揭示嗜乳脂蛋白激活γδ T细胞的“分子钳”机制2025.06.12
- 昌增益课题组揭示蛋白质内源性(顺式)去组装元件调控细菌细胞分裂环(Z环)的动态形成2025.06.09
- 陆剑课题组揭示uORF在动物进化与发育中发挥“翻译减震器”调控作用的新机制2025.06.09
- 张蔚课题组与合作者揭示兰花螳螂程序性体色转变的分子机制及生态学意义2025.05.30
- 2025年“全国科技活动周”和“全国科技工作者日”活动2025.05.29
- 李川昀团队与合作者开发细胞通讯分析工具STCase,揭示肿瘤微环境中空间特异性通讯事件2025.05.29
- 伊成器课题组系统阐释假尿嘧啶修饰的功能与治疗应用新范式2025.05.27
- 瞿礼嘉/钟声团队应邀撰写被子植物雌-雄相互作用保证双受精成功综述2025.05.26
- 陆剑课题组与合作者共同揭示uORFs通过精细调节CLOCK蛋白的翻译参与果蝇昼夜节律及睡眠行为的调控机制2025.05.16
- 伊成器团队发展痕量样品m6A修饰定量测序方法2025.05.08
- 白洋团队联合多位顶尖科学家系统解析根际微生物组调控水稻分蘖的功能与机制2025.04.24
- 瞿礼嘉/钟声课题组发现植物传粉过程中“未雨绸缪”的“两步授粉”备份新机制,为作物逆境下的育性优化提供新思路2025.04.15
- 曾虎课题组应邀撰写综述:空间组学技术的研究进展2025.04.06
- 张迪课题组受邀综述蛋白质L-乳酰化的研究进展2025.04.04
- 苏晓东课题组揭示短序列锚定元件AE在DNA与蛋白质结合中的重要作用2025.03.31
- 魏文胜团队实现人类肿瘤免疫调控网络的单碱基精度解析2025.03.21
- 白洋团队构建全球首个作物根际"细菌+病毒"基因组数据库2025.03.13
- 季雄团队揭示RNA聚合酶亚基RPB7偶联磷酸酶CTDP1稳定Pol II并介导转录再起始2025.03.05
- 何爱彬团队利用全景单细胞组蛋白修饰实现胚胎发育谱系追踪2025.03.04
- 赵进东课题组揭示蓝细菌藻胆体与光系统II结合的新分子机制2025.02.17
- 高歌课题组提出面向大规模异质性空间转录组学切片的表征与解析新方法2025.02.12
- 周岳课题组揭示拟南芥雄性生殖细胞发育过程中染色质三维结构的动态变化过程及其重要作用2025.02.12
- 魏文胜团队发布新一代线粒体碱基编辑器助力建立疾病动物模型2025.01.23
- 贺新强课题组揭示木质部管状分子发育的microRNA调控网络2025.01.19
- 周岳课题组在Genome Biology发文揭示了拟南芥中启动子空间调控模式和喷泉结构形成机制2025.01.02
- 高歌课题组提出人类转录调控元件建模与相关非编码变异功能解析方法2025.01.02
- 魏文胜团队利用碱基编辑器筛选绘制DNA损伤应答功能元件图谱2024.12.16
- 秦跟基课题组揭示弱光下种子萌发调控新机制2024.12.05
- 刘启昆课题组开发了全新的植物细胞谱系追踪工具2024.11.26
- 周岳课题组揭示植物首个三维基因组结构蛋白及其调控机制2024.11.22
- 周岳课题组揭示植物特有的PWWP结构域蛋白调控基因表达的分子机制2024.11.22
- 国家重点实验室陈雪梅教授获得2024年度“求是杰出科学家奖”2024.11.08
- 秦跟基课题组应邀撰写品牌综述“Tansley insight”总结TCP转录因子在细胞器、细胞和器官命运决定中的重要功能2024.10.28
- 郑晓峰课组揭示USP1-ATF4-CD98hc调控ENKTL淋巴瘤患者耐药的新机制2024.09.30
- 肖俊宇课题组揭示IgM–CD5L复合物的分子机制2024.09.30
- 陆剑课题组揭示密码子使用偏好性对翻译调控的影响2024.09.30
- 魏文胜团队实现蛋白质组中丝氨酸、苏氨酸和酪氨酸位点的功能解析2024.09.24
- 王继纵/邓兴旺课题组合作解析植物光敏色素phyB光信号转导的机制2024.09.24
- 刘君/杨雪瑞课题组合作揭示m6A-cenRNA调控癌细胞着丝粒稳态的机制2024.09.23
- 伊成器教授荣获2024年“科学探索奖”2024.08.29